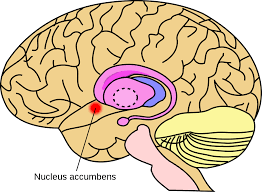
 The nucleus accumbens is a region in the basal forebrain rostral to the preoptic area of the hypothalamus.
The nucleus accumbens is a region in the basal forebrain rostral to the preoptic area of the hypothalamus.
The nucleus accumbens and the olfactory tubercle collectively form the ventral striatum.
The ventral striatum and dorsal striatum collectively form the striatum, which is the main component of the basal ganglia.
Dopaminergic neurons of the mesolimbic pathway project onto the GABAergic medium spiny neurons of the nucleus accumbens and olfactory tubercle.
Each cerebral hemisphere has its own nucleus accumbens.
The nucleus accumbens can be divided into two structures: the nucleus accumbens core and the nucleus accumbens shell.
The nucleus accumbens is an aggregate of neurons with an outer shell and an inner core.
The NA core, shell and neuron subpopulations within each region (D1-type vs D2-type medium spiny neurons) are responsible for different cognitive functions.
The nucleus accumbens has a significant role in the cognitive processing of motivation, aversion, reward, pleasure, positive reinforcement, and reinforcement learning.
The nucleus accumbens is a formation of neurons that is important in reward pathway activation—such as the release of dopamine in investigating response to novel or exciting stimuli.
Part of the nucleus accumbens core is centrally involved in the induction of slow-wave sleep.
The nucleus accumbens plays a lesser role in processing fear, impulsivity, and the placebo effect.
The nucleus accumbens is involved in the encoding of new motor programs.
Major glutamatergic inputs to the nucleus accumbens include: prefrontal cortex, particularly its prelimbic cortex and infralimbic cortex, basolateral amygdala, ventral hippocampus, thalamic nuclei; specifically the midline thalamic nuclei and intralaminar nuclei of the thalamus, and glutamatergic projections from the ventral tegmental area (VTA).
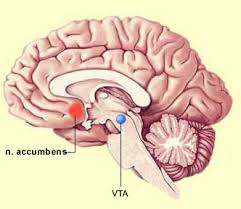
The NA receives dopaminergic inputs from the ventral tegmental area, which connect via the mesolimbic pathway.
It is one part of a cortico-basal ganglia-thalamo-cortical loop.
Dopaminergic inputs from the VTA modulate the activity of GABAergic neurons within the nucleus accumbens.
Such nucleus accumbens neurons are activated directly/: indirectly by euphoriant drugs: amphetamine, opiates, and by participating in rewarding experiences such as sex, music, exercise.
The nucleus accumbens mediates reward, motivation, and task salience.
The majority of reward perceptions involve neuronal activation in the nucleus accumbens, including sex, recreational drugs, and food.
Depolarizations of cells in the nucleus accumbens correlates with positivity of the neurons of the hippocampus, making them more excitable.
The nucleus accumbens receives histaminergic projections from the tuberomammillary nucleus.
The tuberomammillary nucleus is the only source of histamine neurons in the brain.
The nucleus accumbens sends axonal projections to the basal ganglia and the ventral analog of the globus pallidus, known as the ventral pallidum (VP).
The ventral pallidum, in turn, projects to the medial dorsal nucleus of the dorsal thalamus, which projects to the prefrontal cortex as well as the striatum, has efferents to the tail of the ventral tegmental area, substantia nigra, and the reticular formation of the pons.
The nucleus accumbens shell (NAcc shell) is a substructure of the nucleus accumbens.
The shell and core together form the entire nucleus accumbens.
The NA shell is the outer region of the nucleus accumbens.
The NA shell is considered to be part of the extended amygdala, located at its rostral pole.
NA neurons are mostly medium spiny neurons containing mainly D1-type or D2-type dopamine receptors.
A subpopulation of medium spiny neurons contain both D1-type and D2-type receptors, and these mixed-type neurons are mostly confined to the NAcc shell.
NAcc shell neurons,compared to the core, have a lower density of dendritic spines, less terminal segments, and less branch segments than those in the core.
Shell neurons project to the ventral pallidum as well as the ventral tegmental area and to extensive areas in the hypothalamus and extended amygdala.
The shell of the nucleus accumbens is involved in the cognitive processing of reward, and liking reactions to certain pleasurable stimuli, motivational salience, and positive reinforcement.
NAcc shell mediates conditioned stimulus and modifies operant behavior.
NAcc medial shell has a pleasure center which is responsible for the pleasurable or liking component of some intrinsic rewards.
Addictive drugs have a larger effect on dopamine release in the shell than in the core.
The nucleus accumbens core (NAcc core) is the inner substructure of the nucleus accumbens.
The nucleus accumbens core (NAcc core) is located in the nucleus accumbens core is part of the ventral striatum, located within the basal ganglia.
The core of the NAcc is made up mainly of medium spiny neurons containing mainly D1-type or D2-type dopamine receptors.
The D1-type medium spiny neurons mediate reward-related cognitive processes.
D2-type medium spiny neurons mediate aversion-related cognition.
Core neurons as compared to the neurons in the shell, have an increased density of dendritic spines, branch segments, and terminal segments.
Core neurons project to other sub-cortical areas such as the globus pallidus and the substantia nigra.
GABA is one of the main neurotransmitters in the NAcc,
and GABA receptors are also abundant.
The nucleus accumbens core is involved in the cognitive processing of motor function related to reward and reinforcement and the regulation of slow-wave sleep.
The core encodes new motor programs which facilitate the acquisition of a given reward in the future.
The indirect pathway , D2 neurons in the NAcc core which co-express adenosine A2A receptors activation promote slow-wave sleep.
The NAcc core mediates a classically conditioned stimulus modifies operant behavior.
Approximately 95% of neurons in the NAcc are GABAergic medium spiny neurons (MSNs).
GABAergic medium spiny neurons (MSNs) which primarily express either D1-type or D2-type receptors.
About 1–2% of the remaining NAcc neuronal types are large spiny cholinergic interneurons and another 1–2% are GABAergic interneurons.
GABAergic medium spiny neurons in the core have an increased density of dendritic spines, branch segments, and terminal segments compared to the shell.
The neurons of the core project to other sub-cortical areas such as the globus pallidus and the substantia nigra: These neurons are also the main projection or output neurons of the nucleus accumbens.
GABA is one of the main neurotransmitters in the NAcc, and it has many GABA receptors.
The neurotransmitters, neuromodulators, and hormones that signal through receptors within the nucleus accumbens include:
Dopamine, Phenethylamine and tyramine, glucocorticoids, GABA, Glutamate, Serotonin (5-HT)
Dopamine is released into the nucleus accumbens following exposure to rewarding stimuli, and recreational drugs: amphetamines, cocaine, nicotine and morphine.
Phenethylamine and tyramine are trace amines synthesized in neurons expressing the aromatic amino acid hydroxylase (AADC) enzyme, which includes all dopaminergic neurons.
Both compounds function as dopaminergic neuromodulators which regulate the reuptake and release of dopamine into the Nacc.
Glucocorticoid receptors are the only corticosteroid receptors in the nucleus accumbens shell.
GABAA receptors in the NAcc shell have inhibitory control on turning behavior influenced by dopamine, and GABAB receptors have inhibitory control over turning behavior mediated by acetylcholine.
The local blockade of glutamatergic NMDA receptors in the NAcc core impair spatial learning
Serotonin (5-HT) synapses have a greater number of synaptic contacts in the NAcc shell than in the core, along
with larger and thicker, large dense core vesicles than their counterparts in the core.
The nucleus accumbens, is one part of the reward system.
It plays an important role in processing rewarding stimuli, reinforcing stimuli, such as food and water, and rewards and reinforces addictive drugs, sex, and exercise.
The response of neurons in the nucleus accumbens to the reward sucrose is inhibition.
Reducing the excitability of neurons in the nucleus accumbens is rewarding.
The blood oxygen level dependent signal in the nucleus accumbens is increased during the perception of pleasant, emotionally arousing pictures and during mental imagery of pleasant, emotional scenes.
There is an abundance of NAcc inputs from limbic regions and strong NAcc outputs to motor regions, suggesting the NA has an interface between the limbic and motor system.
The nucleus accumbens is causally related to the experience of pleasure.
fMRIs show increased brain activity in the nucleus accumbens and adjacent caudate nucleus, proportionate to the degree to which the women find photos of infants “cute”.
Medium spiny neurons in the nucleus accumbens are involved in reward, whereas the activation of D2-type medium spiny neurons in the nucleus accumbens promotes aversion.
Addiction from chronic drug use involves alterations in gene expression in the mesocorticolimbic projection.
Transcription factors that produce these alterations in gene expression are ΔFosB, cyclic adenosine monophosphate (cAMP) response element binding protein (CREB), and nuclear factor kappa B (NFκB).
ΔFosB is the most significant gene transcription factor in addiction since its overexpression in the nucleus accumbens is necessary and sufficient for many of the neural adaptations and behavioral effects that are seen.
ΔFosB overexpression has been implicated in addictions to alcohol, cannabinoids, cocaine, methylphenidate, nicotine, opioids, phencyclidine, propofol, and substituted amphetamines, among others.
Increases in nucleus accumbens ΔJunD expression can reduce or, with a large increase, even block most of the neural alterations seen in chronic drug abuse by the alterations mediated by ΔFosB.
ΔFosB also plays an important role in regulating behavioral responses to natural rewards, such as palatable food, sex, and exercise.
Natural rewards (drugs of abuse), induce ΔFosB in the nucleus accumbens, and chronic acquisition of these rewards can result in a similar pathological addictive state through ΔFosB overexpression.
ΔFosB is the key transcription factor involved in addictions to natural rewards as well.
ΔFosB in the nucleus accumbens is critical for the reinforcing effects of sexual reward.
Psychostimulants and sexual behavior act on similar biomolecular mechanisms to induce ΔFosB in the nucleus accumbens and possess cross-sensitization effects that are mediated through ΔFosB.
Similar to drug rewards, non-drug rewards also increase the level of extracellular dopamine in the NAcc shell.
Drug-induced dopamine release in the NAcc shell and NAcc core is usually not prone to the development of drug tolerance.
Repeated exposure to drugs that induce dopamine release in the NAcc shell and core typically results in sensitization, with the amount of dopamine that is released in the NAcc from future drug exposure increases as a result of repeated drug exposure.
The sensitization of dopamine release in the NAcc shell following repeated drug exposure serves to strengthen stimulus-drug associations, providing classical conditioning that occurs when drug use is repeatedly paired with environmental stimuli.
These classically conditioned environmental stimuli often become cues which function as secondary reinforcers of drug use.
Contrary to drug effects the release of dopamine in the NAcc shell by many types of rewarding non-drug stimuli typically undergoes habituation following repeated exposure.
The amount of dopamine that is released from future exposure to a rewarding non-drug stimulus normally decreases as a result of repeated exposure to that stimulus.
Deep brain stimulation of the nucleus accumbens was successful in decreasing depression symptoms in 50% of patients who did not respond to other treatments such as electroconvulsive therapy.
It is a target to treat refractory obsessive-compulsive disorder, addiction, mental illness will
radiofrequency ablation of the nucleus accumbens, but results are inconclusive and controversial.
The NAcc activates in the anticipation of effectiveness of a drug when given a placebo, indicating a its contributing role in the placebo effect.