 The complement system is composed of greater than 50 proteins that act in concert to generate products that have both immunoprotective and proinflammatory properties.
The complement system is composed of greater than 50 proteins that act in concert to generate products that have both immunoprotective and proinflammatory properties.
A key component of innate immunity and the regulator of tissue homeostasis.
Complement system, a part of the innate immune system involved in defense against foreign pathogens, clearance of cellular debris, and handling of immune complexes.
Complement is a critical component of the immune system, functioning as a major effector of innate immunity and serving as a bridge to adaptive immune responses.
Its significance lies in several key roles: rapid recognition and elimination of pathogens via opsonization, direct lytic killing through the membrane attack complex, and amplification of inflammation by generating anaphylatoxins such as C3a and C5a, which recruit and activate leukocytes.
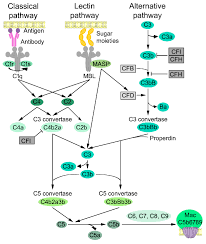
Complement activation occurs through three main pathways—classical, lectin, and alternative; all converging at C3, leading to a cascade that tags pathogens for phagocytosis and triggers cell lysis.
Beyond pathogen defense, complement is essential for homeostasis, including clearance of apoptotic cells and immune complexes, and maintenance of self-tolerance through regulatory proteins that prevent damage to host tissues.
Deficiency or dysregulation of complement components increases susceptibility to infections and contributes to autoimmune and inflammatory diseases, such as systemic lupus erythematosus, atypical hemolytic uremic syndrome, and age-related macular degeneration.
Recent evidence also highlights intracellular complement activity, influencing cell survival, metabolism, and adaptive immune cell function, particularly T cell responses.
Found in greatest concentration in the plasma.
More than 50 soluble and membrane-bound proteins form the complement system, providing innate defense against microbes and mediating inflammatory responses.
Activated via three different pathways: classical, alternative, and lectin.
Importantly, the alternative pathway of complement serves as an amplification loop for the lectin and classical pathways, accounting for roughly 80% of complement activation products.
The complement system is activated by immune complexes as in the classic pathway, or microbial mannose as in the lectin pathway.
The classic and lectin pathways are augmented by the alternative pathway, and which complement factors B and D promote the generation of alternative C5 convertase, C3b.
The alternative pathway, view the action of complement factor P, perpetually promotes the generation of alternative C3b converttase and C3b convertase, resulting in a brisk activation of the complement system for host defense.
The alternative complement system is regulated by a multitude of regulators that include plasma proteins and membrane proteins.
Each pathway leads to the formation of the complement protein C3 convertase and confirm converge on a common terminal pathway, which leads to the formation of the membrane attack complex (MAC), resulting in red blood cell lysis.
Normally CD 55 and CD 59 on red blood cell surfaces act to regulate complement activation.
CD 55 regulates the formation instability of C3 and C5 convertases, and CD 59 regulates terminal complement by blocking the inflammation of the MAC and the insertion of C9 into the lipid bilayer.
On the surface of blood cells and other cells, glycophosphatidylinositol anchored CD55 and CD59 are involved in the regulation of the alternative complement pathway.
In PNH, somatic mutation of the PIGA gene in a hematopoetic stem cell causes deficiency of a glycophosphatidylinositol in its progeny cells: as a result, excessive activation of the complement system occurs on the surface of the affected blood cells, resulting in intravascular hemolysis and thrombosis.
With PNH there is constant, low level complemented activity through the alternative pathway leading to chronic intravascular hemolysis.
Complement amplifying events such as infection, surgery, pregnancy, vaccination, or other inflammatory triggers can lead to complement activation to any of the proximal pathways, causing severe paroxysmal events.
All activating pathways converge to complement component 3-mediated amplification loop by pathway-specific C3 convertases.
The classical pathway is activated by an antigen-antibody immune complex or C-reactive protein, and the lectin pathway is activated directly by mannose-containing bacterial surfaces.
The classical pathway is mainly activated by antibody-antigen complexes recognized via complement component C1q.
The liver is the major site for the synthesis of most complement molecules.
Besides hepatocytes, other cell types, including monocytes, macrophages, endothelial cells, fibroblasts, and adipocytes, can be local sources of complement proteins such as C1q, C3, and C529.
Among them, both activating molecules (C3, C4, C9, and factor B) and negative regulators (C1 inhibitor and C4BP) are up-regulated during an acute-phase reaction, which underlines the relevance of activating balanced complement-mediated responses.
Among antibody isotypes, IgM is the most effective in activating complement.
Activation of complement with the four subclasses of IgGs varies as a function of steric hindrance by the Fab arms in the approach of C1q to the IgG CH2 sites (IgG3>IgG1>IgG2>IgG4).
The system functions by innate and adaptive immunity for defense of microbes.
Activation results formation of a number of components that increase vasculature permeability, chemotaxis and opsonization.
Present in inactive forms in plasma with numeration of C1-C9.
Many complement proteins are activated to become proteolytic enzymes that degrade therapy complement proteins forming a cascade effect.
Plays a role in acute inflammatory response associated with ischemia and reperfusion injury.
C3a and C5a which are anaphylatoxins, and C5b-9 membrane attack complex formed during complement activation promote tissue injury by increasing vascular permeability, activates endothelium, activates hemolysis, induces cell lysis, and induces apoptosis.