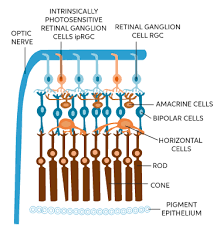
 Retinal ganglion cells are specialized neurons located in the retina of the eye.
Retinal ganglion cells are specialized neurons located in the retina of the eye.
They play a key role in the process of vision by transmitting visual information from the retina to the brain.
They receive signals from the outer retina (photoreceptors and bipolar cells) and transmit this information through their axons, which form the optic nerve that connects the retina to the brain.
There are different types of retinal ganglion cells, each responding to different features of the visual environment, such as contrast, color, orientation, and motion.
Some retinal ganglion cells are sensitive to the overall brightness or darkness of the visual image, while others are involved in detecting specific shapes and patterns.
Retinal ganglion cells are involved in visual perception and can be affected in various visual disorders, such as glaucoma and optic neuropathy.
A retinal ganglion cell (RGC) is a type of neuron located near the inner surface, or the ganglion cell layer of the retina of the eye.
RGCs receive visual information from photoreceptors via two intermediate neuron types: bipolar cells and retina amacrine cells.
Retina amacrine cells, create functional subunits within the ganglion cell layer and making it so that ganglion cells can observe a small dot moving a small distance.
Retinal ganglion cells transmit image-forming and non-image forming visual information from the retina in the form of action potential to several regions in the thalamus, hypothalamus, and mesencephalon, or midbrain.
Retinal ganglion cells have a long axon that extend into the brain, and form the optic nerve, optic chiasm, and optic tract.
A small percentage of retinal ganglion cells are photosensitive, and their axons form the retinohypothalamic tract and contribute to circadian rhythms and pupillary light reflex resizing.
Photosensitive retinal ganglion cells, are involved in modulating the circadian rhythm because of the expression of melanopsin, which absorbs light in the blue part (around 480 nm).
There are about 0.7 to 1.5 million retinal ganglion cells in the human retina.
There are about 4.6 million cone cells and 92 million rod cells, or 96.6 million photoreceptors per retina,
On average each retinal ganglion cell receives inputs from about 100 rods and cones.
These numbers vary among individuals and as a function of retinal location.
In the fovea, a single ganglion cell will communicate with as few as five photoreceptors.
In the extreme periphery of the retina, a single ganglion cell will receive information from many thousands of photoreceptors.
Retinal ganglion cells spontaneously fire action potentials at a base rate, while at rest.
Tbere are generally three classes of RGC ganglions.
W-ganglion: small, 40% of total RHCs, have broad fields in retina, and excitation from rods.
It detects direction of movement anywhere in the field.
X-ganglion: is of medium diameter, and makes up 55% of total, small field, and color vision.
Has sustained response.
Y- ganglion: largest, 5%, very broad dendritic field, respond to rapid eye movement or rapid change in light intensity.
There are at least five main classes of retinal ganglion cells:
Midget cell (parvocellular, or P pathway; P cells)
Parasol cell (magnocellular, or M pathway; M cells)
Bistratified cell (koniocellular, or K pathway)
Photosensitive ganglion cells
Other ganglion cells projecting to the superior colliculus for eye movements (saccades)
P-type retinal ganglion cells project to the parvocellular layers of the lateral geniculate nucleus.
These cells are known as midget retinal ganglion cells. ,
P cells have small sizes of their dendritic trees and cell bodies.
About 80% of all retinal ganglion cells are midget cells in the parvocellular pathway.
Midget cells in the parvocellular pathway receive inputs from relatively few rods and cones.
Midget cells have slow conduction velocity, and respond to changes in color but respond only weakly to changes in contrast.
Midget cells project to the magnocellular layers of the lateral geniculate nucleus.
M-pathway cells are known as parasol retinal ganglion cells, based on the large sizes of their dendritic trees and cell bodies.
About 10% of all retinal ganglion cells are parasol cells, and these cells are part of the magnocellular pathway.
They receive inputs from relatively many rods and cones, have fast conduction velocity, and can respond to low-contrast stimuli, but are not very sensitive to changes in color.
There are more transient temporal responses in this pathway, compared to the P-type, and this retinal pathway is largely color blind.
BiK-type retinal ganglion cells project to the koniocellular layers of the lateral geniculate nucleus.
About 10% of all retinal ganglion cells are bistratified cells, and these cells go through the koniocellular pathway.
They receive inputs from intermediate numbers of rods and cones, and
may be involved in color vision.
Photosensitive ganglion cells, including the giant retinal ganglion cells, contain their own photopigment, melanopsin, which makes them respond directly to light even in the absence of rods and cones.
Photosensitive ganglion cells project to the suprachiasmatic nucleus (SCN) via the retinohypothalamic tract for setting and maintaining circadian rhythms.
Different retinal ganglion cells project to the lateral geniculate nucleus (LGN) with connections to the Edinger-Westphal nucleus (EW), for control of the pupillary light reflex, and giant retinal ganglion cells.
Degeneration of axons of the retinal ganglion cells of the optic nerve is a hallmark of glaucoma.
Axons from the RGCs grow and extend towards the optic disc, where they exit the eye.
RGCs exit the retinal ganglion cell layer through the optic disc.
RGCs grow along glial cell end feet in the optic nerve.
RGC axons traveling to the contralateral optic tract need to cross.
Once out of the optic chiasm, RGCs extend dorsocaudally to the optic tract, guiding them to the superior colliculus and lateral geniculate nucleus.
The axons of retinal ganglion cells are not myelinated where they pass through the retina, but parts of axons that are beyond the retina, are myelinated.