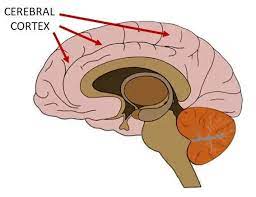
 The cerebral cortex is the outer layer of neural tissue of the cerebrum of the brain.
The cerebral cortex is the outer layer of neural tissue of the cerebrum of the brain.
The cerebral cortex mostly consists of the six-layered neocortex, with just 10% consisting of allocortex.
The cerebral cortex can be classified into two types, the large area of neocortex which has six cell layers, and the much smaller area of allocortex that has three or four layers:
The cerebral cortex is separated into two cortices, by the longitudinal fissure that divides the cerebrum into the left and right cerebral hemispheres.
The two hemispheres are joined beneath the cortex by the corpus callosum.
The cerebral cortex is the largest site of neural integration in the central nervous system.
The cerebral cortex plays a key role in attention, perception, awareness, thought, memory, language, and consciousness, and is part of the brain responsible for cognition.
The cerebral cortex is folded, providing a greater surface area in the confined volume of the cranium.
The cortex folding minimizes brain and cranial volume, and is crucial for the brain circuitry and its functional organization.
When unfolded in the human, each hemispheric cortex has a total surface area of about 0.12 square metres (1.3 sq ft).
The folding is inward away from the surface of the brain, and is also present on the medial surface of each hemisphere within the longitudinal fissure.
In mammals with small brains there is no folding and the cortex is smooth.
A fold or ridge in the cortex is termed a gyrus.
A cortical groove is termed a sulcus.
The cerebral cortex is the outer covering of the surfaces of the cerebral hemispheres and is folded into peaks called gyri, and grooves called sulci.
These surface convolutions appear during fetal development and continue to mature after birth by the process of gyrification.
The majority of the cerebral cortex is not visible from the outside, but buried in the sulci.
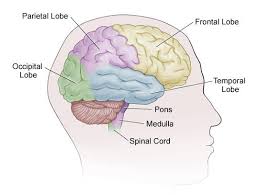
The major sulci and gyri mark the divisions of the cerebrum into the lobes of the brain.
The four major lobes are the frontal, parietal, occipital and temporal lobes.
Other lobes are the limbic lobe, and the insular cortex often referred to as the insular lobe.
Each lobe is associated with different functions, such as the frontal lobe’s role in decision-making and problem-solving, and the occipital lobe’s role in visual processing.
The cerebral cortex is the outer layer of the brain, made up of folded gray matter.
It plays a key role in consciousness, perception, memory, language, thought, and consciousness.
The frontal lobe is located at the front of the brain and is commonly associated with executive functions, including decision-making, problem-solving, planning, reasoning, and controlling voluntary movements.
The parietal lobe is located at the top and back of the brain and is responsible for processing sensory information from the body, such as touch, pressure, pain, temperature, and spatial awareness.
The temporal lobe is located on the sides of the brain and is mainly involved in the processing of auditory and visual information.
It is also involved in memory and language comprehension.
The occipital lobe is located at the back of the brain and is mainly responsible for processing visual information from the eyes.
Overall, the functions of each lobe are interconnected, and multiple areas of the brain work together to process information and carry out complex tasks.
There are between 14 and 16 billion neurons in the human cerebral cortex.
The cortical neurons are organized into horizontal cortical layers, and radially into cortical columns and minicolumns.
Cortical areas have specific functions such as movement in the motor cortex, and sight in the visual cortex.
Visual cortex is located in the occipital lobe.
The cerebral cortex is between two and three or four millimetres thick, and makes up 40 per cent of the brain’s mass.
90 per cent of the cerebral cortex is the six-layered neocortex with the other 10 per cent made up of allocortex.
The neocortex is separable into different regions of cortex and include the motor cortex and visual cortex.
About two thirds of the cortical surface is buried in the sulci and the insular cortex is completely hidden.
The cortex is thickest over the top of a gyrus and thinnest at the bottom of a sulcus.
The larger sulci and gyri mark the divisions of the cortex of the cerebrum into the lobes of the brain.
There are four main lobes: the frontal lobe, parietal lobe, temporal lobe, and occipital lobe.
The insular cortex is often included as the insular lobe.
The limbic lobe is a rim of cortex on the medial side of each hemisphere and is also often included.
There are also three lobules of the brain described: the paracentral lobule, the superior parietal lobule, and the inferior parietal lobule.
Magnetic resonance imaging of the brain (MRI) allows for measurement of the thickness of the human cerebral cortex.
The thickness of different cortical areas varies: sensory cortex is thinner than motor cortex.
There may be some positive association between the cortical thickness and intelligence.
The somatosensory cortex is thicker in migraine patients.
Dyslexia is a genetic disorder of the cerebral cortex, whereby decreased folding in certain areas results in a microgyrus, where there are four layers instead of six,
The neocortex is formed of six layers, numbered I to VI, from the outermost layer I – near to the pia mater, to the innermost layer VI – near to the underlying white matter.
Each cortical layer has a characteristic distribution of different neurons and their connections with other cortical and subcortical regions.
There are direct connections between different cortical areas and indirect connections via the thalamus.
Brodmann established that the mammalian neocortex is consistently divided into six layers.
Layer I is the molecular layer, and contains few scattered neurons, including GABAergic rosehip neurons.
It consists largely of extensions of apical dendritic tufts of pyramidal neurons and horizontally oriented axons, as well as glial cells.
Inputs to the apical tufts are thought to be crucial for the feedback interactions in the cerebral cortex involved in associative learning and attention.
Layer I across the cerebral cortex mantle receives substantial input from matrix or M-type thalamus cells.
Layer II is the external granular layer, which contains small pyramidal neurons and numerous stellate neurons.
Layer III, the external pyramidal layer, contains predominantly small and medium-size pyramidal neurons, as well as non-pyramidal neurons with vertically oriented intracortical axons.
Layers I through III are the main target of interhemispheric corticocortical afferents, and layer III is the principal source of corticocortical efferents.
Layer IV, the internal granular layer, contains different types of stellate and pyramidal cells, and is the main target of thalamocortical afferents from thalamus type C neurons as well as intra-hemispheric corticocortical afferents.
The layers above layer IV are also referred to as supragranular layers (layers I-III), whereas the layers below are referred to as infragranular layers (layers V and VI).
Layer V, the internal pyramidal layer, contains large pyramidal neurons.
Axons from level V leave the cortex and connect with subcortical structures including the basal ganglia.
In the primary motor cortex of the frontal lobe, layer V contains giant pyramidal cells called Betz cells, whose axons travel through the internal capsule, the brain stem, and the spinal cord forming the corticospinal tract, which is the main pathway for voluntary motor control.
Layer VI is the polymorphic or multiform layer, contains few large pyramidal neurons and many small spindle-like pyramidal and multiform neurons.
Layer VI sends efferent fibers to the thalamus, establishing a very precise reciprocal interconnection between the cortex and the thalamus.
Layer VI sends efferent fibers to the thalamus, and these connections are both excitatory and inhibitory.
Neurons send excitatory fibers to neurons in the thalamus and also send collaterals to the thalamic reticular nucleus that inhibit these same thalamus neurons or ones adjacent to them, providing the brainstem with adjustable control.
There are characteristic connections between different layers and neuronal types, which span all the thickness of the cortex.
These cortical microcircuits are grouped into cortical columns and minicolumns.
It is suggested that the minicolumns are the basic functional units of the cortex.
Blood supply to the cerebral cortex is part of the cerebral circulation.
Arterial blood carries oxygen, glucose, and other nutrients to the cortex.
Cerebral veins drain the deoxygenated blood, and metabolic wastes including carbon dioxide, back to the heart.
The main arteries supplying the cortex are the anterior cerebral artery, the middle cerebral artery, and the posterior cerebral artery.
The anterior cerebral artery supplies the anterior portions of the brain, including most of the frontal lobe.
The middle cerebral artery supplies the parietal lobes, temporal lobes, and parts of the occipital lobes.
The middle cerebral artery splits into two branches to supply the left and right hemisphere, where they branch further.
The posterior cerebral artery supplies the occipital lobes.
The circle of Willis is the main blood system that deals with blood supply in the cerebrum and cerebral cortex.
The cerebral cortex is composed of a heterogenous population of cells.
The majority of these cells are derived from radial glia migration that form the different cell types of the neocortex.
This process of neurogenesis regulates lamination to form the different layers of the cortex.
There is an an inside-out topography in the cortex with younger neurons in superficial layers and older neurons in deeper layers.
At birth there are very few dendrites present on the cortical neuron’s cell body, and the axon is undeveloped.
During the first year of life the dendrites become dramatically increased in number, such that they can accommodate up to a hundred thousand synaptic connections with other neurons.
The axon can develop to extend a long way from the cell body.
Cortical development between 26 and 39 week gestational age, and
The layered structure of the mature cerebral cortex is formed during development.
Rapid expansion of the cortical surface area is regulated by the amount of self-renewal of radial glial cells and is partly regulated by FGF and Notch genes.
During the period of cortical neurogenesis and layer formation, many the process of gyrification begins, which generates the characteristic folds of the cerebral cortex.
The cerebral cortex is connected to subcortical structures such as the thalamus and the basal ganglia, sending information to them along efferent connections and receiving information from them via afferent connections.
Most sensory information is routed to the cerebral cortex via the thalamus.
Olfactory information passes through the olfactory bulb to the olfactory cortex.
The majority of connections are from one area of the cortex to another, rather than from subcortical areas.
The whole of the cerebral cortex was divided into 52 different areas in an early presentation Brodmann.
These areas known as Brodmann areas, are based on their cytoarchitecture but also relate to various functions.
Tee cerebral cortex is typically described as comprising three parts: sensory, motor, and association areas.
The sensory areas are the cortical areas receive and process information from the senses.
Parts of the cortex that receive sensory inputs from the thalamus are called primary sensory areas: senses of vision, hearing, and touch are served by the primary visual cortex, primary auditory cortex and primary somatosensory cortex, respectively.
The two hemispheres receive information from the opposite side of the body: the right primary somatosensory cortex receives information from the left limbs, and the right visual cortex receives information from the left visual field.
The organization of sensory maps in the cortex reflects that of the corresponding sensing organ.
A topographic map exists of neighboring points: the primary visual cortex correspond to neighboring points in the retina; a tonotopic map in the primary auditory cortex and a somatotopic map in the primary sensory cortex; topographic map of the posterior central gyrus has been illustrated as a deformed human representation, the somatosensory homunculus, where the size of different body parts reflects the relative density of their innervation.
Areas with much sensory innervation, such as the fingertips and the lips, require more cortical area to process finer sensation.
The motor areas are located in both hemispheres of the cortex.
The right half of the motor area controls the left side of the body, and vice versa.
Two areas of the cortex are commonly referred to as motor:
Primary motor cortex, which executes voluntary movements
Supplementary motor areas and premotor cortex, which select voluntary movements.
The posterior parietal cortex guides voluntary movements in space.
The dorsolateral prefrontal cortex decides which voluntary movements to make according to higher-order instructions, rules, and self-generated thoughts.
Just underneath the cerebral cortex are interconnected subcortical masses of grey matter, the basal ganglia (or nuclei).
The basal ganglia receive input from the substantia nigra of the midbrain and motor areas of the cerebral cortex, and send signals back to both of these locations.
The basal ganglia are involved in motor control, and are found lateral to the thalamus.
The main components of the basal ganglia are the caudate nucleus, the putamen, the globus pallidus, the substantia nigra, the nucleus accumbens, and the subthalamic nucleus.
The putamen and globus pallidus are also collectively known as the lentiform nucleus, because together they form a lens-shaped body.
The putamen and caudate nucleus are also collectively called the corpus striatum after their striped appearance.
Association areas are the parts of the cerebral cortex that do not belong to the primary regions.
Association areas of the cerebral cortex function to produce a perceptual experiences of the world, to enable interactions, and support abstract thinking and language.
The parietal, temporal, and occipital lobes – all located in the posterior part of the cortex – integrate sensory information and information stored in memory.
The frontal lobe or prefrontal association complex is involved in planning actions and movement, as well as abstract thought.
The ssociation networks are particularly important to language function.
In the past it was theorized that language abilities are localized in Broca’s area in areas of the left inferior frontal gyrus, BA44 and BA45, for language expression and in Wernicke’s area BA22, for language reception.
It has been been shown to occur in areas other than just those structures around the lateral sulcus, including the frontal lobe, basal ganglia, cerebellum, and pons.
Cortical stimulation mapping is an invasive procedure that involves placing electrodes directly onto the exposed brain in order to localise the functions of specific areas of the cortex.
It is used in clinical and therapeutic applications including pre-surgical mapping.
A number of genetic mutations that can cause a wide range of genetic disorders of the cerebral cortex, including microcephaly, schizencephaly and types of lissencephaly.
Chromosome abnormalities can also result causing a number of neurodevelopmental disorders such as fragile X syndrome and Rett syndrome.
MCPH1 codes for microcephalin, and disorders in this and in ASPM are associated with microcephaly.
Mutations in the gene NBS1 that codes for nibrin can cause Nijmegen breakage syndrome, characterised by microcephaly.
Mutations in EMX2, and COL4A1 are associated with schizencephaly, a condition marked by the absence of large parts of the cerebral hemispheres.